Die aerobe Atmung benötigt Sauerstoff (O2), um ATP zu erzeugen. Obwohl Kohlenhydrate, Fette und Proteine als Reaktanten verbraucht werden, ist die aerobe Atmung die bevorzugte Methode des Pyruvatabbaus in der Glykolyse und erfordert Pyruvat zu den Mitochondrien, um vollständig durch den Zitronensäurezyklus oxidiert zu werden. Die Produkte dieses Prozesses sind Kohlendioxid und Wasser, und die übertragene Energie wird verwendet, um Bindungen in ADP zu brechen, um eine dritte Phosphatgruppe hinzuzufügen, um ATP (Adenosintriphosphat), durch Phosphorylierung auf Substratebene, NADH und FADH2 zu bilden
| Vereinfachte Reaktion: | C6H12O6 (s) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + Wärme |
| ΔG = -2880 kJ pro Mol C6H12O6 |
Das negative ΔG zeigt an, dass die Reaktion spontan ablaufen kann.
Das Potential von NADH und FADH2 wird durch eine Elektronentransportkette mit Sauerstoff und Protonen (Wasserstoff) als „terminalen Elektronenakzeptoren“ in mehr ATP umgewandelt. Der größte Teil des ATP, das bei der aeroben Zellatmung entsteht, wird durch oxidative Phosphorylierung gebildet. Die Energie des freigesetzten O2 wird genutzt, um ein chemiosmotisches Potential zu erzeugen, indem Protonen über eine Membran gepumpt werden. Dieses Potential wird dann verwendet, um die ATP-Synthase anzutreiben und ATP aus ADP und einer Phosphatgruppe zu produzieren. In Biologie-Lehrbüchern wird oft angegeben, dass pro oxidiertem Glukosemolekül während der Zellatmung 38 ATP-Moleküle hergestellt werden können (2 aus der Glykolyse, 2 aus dem Krebs-Zyklus und etwa 34 aus dem Elektronentransportsystem). Diese maximale Ausbeute wird jedoch aufgrund von Verlusten durch undichte Membranen sowie der Kosten für den Transport von Pyruvat und ADP in die mitochondriale Matrix nie ganz erreicht, und aktuelle Schätzungen liegen bei etwa 29 bis 30 ATP pro Glukose.
Der anaerobe Stoffwechsel ist bis zu 15-mal effizienter als der anaerobe Stoffwechsel (der 2 Moleküle ATP pro 1 Molekül Glukose liefert), da die Doppelbindung in O2 eine höhere Energie hat als andere Doppelbindungen oder Paare von Einfachbindungen in anderen gängigen Molekülen in der Biosphäre. Einige anaerobe Organismen, wie z. B. Methanogene, sind jedoch in der Lage, mit der anaeroben Atmung fortzufahren und mehr ATP zu gewinnen, indem sie andere anorganische Moleküle (nicht Sauerstoff) mit hoher Energie als endgültige Elektronenakzeptoren in der Elektronentransportkette verwenden. Sie teilen den anfänglichen Weg der Glykolyse, aber der aerobe Stoffwechsel wird mit dem Krebszyklus und der oxidativen Phosphorylierung fortgesetzt. Die post-glykolytischen Reaktionen finden bei eukaryotischen Zellen in den Mitochondrien und bei prokaryotischen Zellen im Zytoplasma statt.
Glykolyse
Die Glykolyse ist ein Stoffwechselweg, der in allen lebenden Organismen im Cytosol der Zellen stattfindet. Glykolyse kann wörtlich mit „Zuckerspaltung“ übersetzt werden und findet mit oder ohne Anwesenheit von Sauerstoff statt. Unter aeroben Bedingungen wandelt der Prozess ein Molekül Glukose in zwei Moleküle Brenztraubensäure (Pyruvat) um und erzeugt dabei Energie in Form von zwei Nettomolekülen ATP. Eigentlich werden vier Moleküle ATP pro Glukose produziert, zwei werden jedoch im Rahmen der Vorbereitungsphase verbraucht. Die anfängliche Phosphorylierung der Glukose ist erforderlich, um die Reaktivität zu erhöhen (ihre Stabilität zu verringern), damit das Molekül durch das Enzym Aldolase in zwei Pyruvatmoleküle gespalten werden kann. Während der Pay-off-Phase der Glykolyse werden vier Phosphatgruppen durch Phosphorylierung auf Substratebene auf ADP übertragen, um vier ATP zu bilden, und zwei NADH werden produziert, wenn das Pyruvat oxidiert wird. Die Gesamtreaktion kann so ausgedrückt werden:
Glucose + 2 NAD+ + 2 Pi + 2 ADP → 2 Pyruvat + 2 H+ + 2 NADH + 2 ATP + 2 H+ + 2 H2O + Energie
Ausgehend von Glucose wird 1 ATP verwendet, um ein Phosphat an Glucose zu spenden, um Glucose-6-Phosphat zu erzeugen. Auch Glykogen kann mit Hilfe der Glykogenphosphorylase in Glukose-6-Phosphat umgewandelt werden. Im Energiestoffwechsel wird aus Glukose-6-Phosphat Fruktose-6-Phosphat. Mit Hilfe der Phosphofructokinase wird Fructose-6-Phosphat mit zusätzlichem ATP zu Fructose-1,6-Bisphosphat phosphoryliert. Fructose 1,6-Biphosphat spaltet sich dann in zwei phosphorylierte Moleküle mit drei Kohlenstoffketten, die später zu Pyruvat abgebaut werden.
Oxidative Decarboxylierung von Pyruvat
Pyruvat wird durch den Pyruvat-Dehydrogenase-Komplex (PDC) zu Acetyl-CoA und CO2 oxidiert. Der PDC enthält mehrere Kopien von drei Enzymen und befindet sich in den Mitochondrien von eukaryotischen Zellen und im Cytosol von Prokaryoten. Bei der Umwandlung von Pyruvat zu Acetyl-CoA entstehen ein Molekül NADH und ein Molekül CO2.
Zitronensäurezyklus
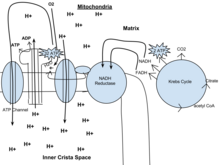
Dieser wird auch als Krebszyklus oder Tricarbonsäurezyklus bezeichnet. Bei Anwesenheit von Sauerstoff wird aus den bei der Glykolyse entstandenen Pyruvatmolekülen Acetyl-CoA hergestellt. Sobald Acetyl-CoA gebildet ist, kann aerobe oder anaerobe Atmung stattfinden. Wenn Sauerstoff vorhanden ist, findet in den Mitochondrien eine aerobe Atmung statt, die zum Krebszyklus führt. Wenn jedoch kein Sauerstoff vorhanden ist, kommt es zur Fermentation des Pyruvatmoleküls. In Gegenwart von Sauerstoff, wenn Acetyl-CoA produziert wird, tritt das Molekül dann in den Zitronensäurezyklus (Krebszyklus) innerhalb der Mitochondrienmatrix ein und wird zu CO2 oxidiert, während gleichzeitig NAD zu NADH reduziert wird. Das NADH kann von der Elektronentransportkette verwendet werden, um im Rahmen der oxidativen Phosphorylierung weiteres ATP zu erzeugen. Um das Äquivalent eines Glukosemoleküls vollständig zu oxidieren, müssen zwei Acetyl-CoA durch den Krebszyklus verstoffwechselt werden. Bei diesem Zyklus entstehen zwei energiearme Abfallprodukte, H2O und CO2.
Der Zitronensäurezyklus ist ein 8-stufiger Prozess, an dem 18 verschiedene Enzyme und Co-Enzyme beteiligt sind. Während des Zyklus entsteht aus Acetyl-CoA (2 Kohlenstoffe) + Oxalacetat (4 Kohlenstoffe) Citrat (6 Kohlenstoffe), das zu einer reaktiveren Form namens Isocitrat (6 Kohlenstoffe) umgewandelt wird. Isocitrat wird zu α-Ketoglutarat (5 Kohlenstoffe), Succinyl-CoA, Succinat, Fumarat, Malat und schließlich Oxalacetat modifiziert.
Der Nettogewinn aus einem Zyklus sind 3 NADH und 1 FADH2 als wasserstoff- (protonen- und elektronen-) tragende Verbindungen und 1 energiereiches GTP, das anschließend zur Herstellung von ATP verwendet werden kann. Die Gesamtausbeute aus 1 Glukosemolekül (2 Pyruvatmoleküle) beträgt also 6 NADH, 2 FADH2 und 2 ATP.
Oxidative Phosphorylierung
In Eukaryoten findet die oxidative Phosphorylierung in den mitochondrialen Cristae statt. Sie umfasst die Elektronentransportkette, die durch die Oxidation des aus dem Krebszyklus stammenden NADH einen Protonengradienten (chemiosmotisches Potential) über die Grenze der inneren Membran aufbaut. ATP wird durch das Enzym ATP-Synthase synthetisiert, wenn der chemiosmotische Gradient genutzt wird, um die Phosphorylierung von ADP anzutreiben. Der Elektronentransfer wird durch die chemische Energie des exogenen Sauerstoffs angetrieben und unter Zugabe von zwei Protonen wird Wasser gebildet.