Respiração aeróbica requer oxigénio (O2) a fim de criar ATP. Embora os hidratos de carbono, gorduras e proteínas sejam consumidos como reagentes, a respiração aeróbica é o método preferido de degradação pirúvica em glicólise, e requer piruvato para as mitocôndrias a fim de ser totalmente oxidado pelo ciclo do ácido cítrico. Os produtos deste processo são dióxido de carbono e água, e a energia transferida é utilizada para quebrar ligações em ADP para adicionar um terceiro grupo de fosfatos para formar ATP (trifosfato de adenosina), por fosforilação ao nível do substrato, NADH e FADH2
| Reacção simplificada: | C6H12O6 (s) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + calor |
O negativo ΔG indica que a reacção pode ocorrer espontaneamente.
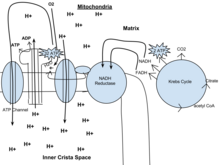
O potencial de NADH e FADH2 é convertido em mais ATP através de uma cadeia de transporte de electrões com oxigénio e prótons (hidrogénio) como os “aceitadores terminais de electrões”. A maior parte do ATP produzido pela respiração celular aeróbica é feita por fosforilação oxidativa. A energia do O2 libertado é utilizada para criar um potencial quimiossimótico através da bombagem de protões através de uma membrana. Este potencial é então utilizado para conduzir a ATP synthase e produzir ATP a partir de ADP e de um grupo fosfato. Os livros de biologia afirmam frequentemente que 38 moléculas de ATP podem ser feitas por molécula de glucose oxidada durante a respiração celular (2 da glicólise, 2 do ciclo de Krebs, e cerca de 34 do sistema de transporte de electrões). No entanto, este rendimento máximo nunca é completamente atingido devido a perdas devidas a fugas de membranas, bem como ao custo de mover o piruvato e o ADP para a matriz mitocondrial, e as estimativas actuais variam cerca de 29 a 30 ATP por glucose.
O metabolismo aeróbio é até 15 vezes mais eficiente do que o metabolismo anaeróbio (que produz 2 moléculas de ATP por 1 molécula de glucose) porque a ligação dupla em O2 é de maior energia do que outras ligações duplas ou pares de ligações simples em outras moléculas comuns na biosfera. No entanto, alguns organismos anaeróbios, tais como os metanogénicos são capazes de continuar com a respiração anaeróbia, produzindo mais ATP através da utilização de outras moléculas inorgânicas (não oxigénio) de alta energia como aceitadores finais de electrões na cadeia de transporte de electrões. Partilham a via inicial da glicólise, mas o metabolismo aeróbio continua com o ciclo de Krebs e a fosforilação oxidativa. As reacções pós-colíticas ocorrem nas mitocôndrias em células eucarióticas, e no citoplasma em células procarióticas.
Glycolysis
glicólise é uma via metabólica que tem lugar no citosol das células em todos os organismos vivos. A glicólise pode ser literalmente traduzida como “divisão do açúcar”, e ocorre com ou sem a presença de oxigénio. Em condições aeróbicas, o processo converte uma molécula de glucose em duas moléculas de piruvato (ácido pirúvico), gerando energia sob a forma de duas moléculas líquidas de ATP. Quatro moléculas de ATP por glicose são efectivamente produzidas, contudo, duas são consumidas como parte da fase preparatória. A fosforilação inicial da glucose é necessária para aumentar a reactividade (diminuir a sua estabilidade) para que a molécula seja clivada em duas moléculas de piruvato pela enzima aldolase. Durante a fase de pay-off da glicólise, quatro grupos de fosfato são transferidos para o ADP por fosforilação a nível de substrato para fazer quatro ATP, e dois NADH são produzidos quando o piruvato é oxidado. A reacção global pode ser expressa desta forma:
Glicose + 2 NAD+ + 2 Pi + 2 ADP → 2 piruvato + 2 H+ + 2 NADH + 2 ATP + 2 H+ + 2 H2O + energia
Início com glicose, 1 ATP é utilizado para doar um fosfato à glicose para produzir glicose 6-fosfato. O glicogénio também pode ser convertido em glicose 6-fosfato com a ajuda de glicogénio fosforilase. Durante o metabolismo energético, a glicose 6-fosfato transforma-se em frutose 6-fosfato. Um ATP adicional é utilizado para fosforilato de frutose 6-fosfato em frutose 1,6-bisfosfato com a ajuda da fosfofructoquinase. A frutose 1,6-bifosfato divide-se depois em duas moléculas fosforiladas com três cadeias de carbono que mais tarde se degradam em piruvato.
Descarboxilação oxidativa do piruvato
O piruvato é oxidado a acetil-CoA e CO2 pelo complexo de piruvato desidrogenase (PDC). O PDC contém múltiplas cópias de três enzimas e está localizado nas mitocôndrias de células eucarióticas e no citosol de procariotas. Na conversão do piruvato em acetil-CoA, forma-se uma molécula de NADH e uma molécula de CO2.
Ciclo do ácido cítrico
Este também é chamado o ciclo de Krebs ou o ciclo do ácido tricarboxílico. Quando o oxigénio está presente, a acetil-CoA é produzido a partir das moléculas pirúvel criadas a partir da glicólise. Uma vez formada a acetil-CoA, pode ocorrer respiração aeróbica ou anaeróbica. Quando o oxigénio está presente, as mitocôndrias serão submetidas à respiração aeróbica, o que leva ao ciclo de Krebs. No entanto, se o oxigénio não estiver presente, ocorrerá a fermentação da molécula pirúvica. Na presença de oxigénio, quando a acetil-CoA é produzida, a molécula entra então no ciclo do ácido cítrico (ciclo de Krebs) dentro da matriz mitocondrial, e é oxidada a CO2, ao mesmo tempo que reduz o NAD a NADH. O NADH pode ser utilizado pela cadeia de transporte de electrões para criar mais ATP como parte da fosforilação oxidativa. Para oxidar completamente o equivalente a uma molécula de glucose, duas acetil-CoA devem ser metabolizadas pelo ciclo de Krebs. Durante este ciclo são criados dois produtos residuais de baixo consumo energético, H2O e CO2.
O ciclo do ácido cítrico é um processo de 8 etapas que envolve 18 enzimas e co-enzimas diferentes. Durante o ciclo, o acetil-CoA (2 carbonos) + oxaloacetato (4 carbonos) produz citrato (6 carbonos), que é rearranjado para uma forma mais reactiva chamada isocitrato (6 carbonos). O isocitrato é modificado para se tornar α-ketoglutarato (5 carbonos), succinil-CoA, succinato, fumarato, malato, e, finalmente, oxaloacetato.
O ganho líquido de um ciclo é de 3 NADH e 1 FADH2 como compostos portadores de hidrogénio (protão mais electrão) e 1 GTP de alta energia, que pode ser subsequentemente utilizado para produzir ATP. Assim, o rendimento total de 1 molécula de glucose (2 moléculas pirúvel) é de 6 NADH, 2 FADH2, e 2 ATP.
Fosforilação oxidativa
Em eucariotas, a fosforilação oxidativa ocorre na cristae mitocondrial. Compreende a cadeia de transporte de electrões que estabelece um gradiente de prótons (potencial quimiosmótico) através do limite da membrana interna, oxidando o NADH produzido a partir do ciclo de Krebs. O ATP é sintetizado pela enzima ATP synthase quando o gradiente quimiossimótico é utilizado para conduzir a fosforilação do ADP. A transferência de electrões é impulsionada pela energia química do oxigénio exógeno e, com a adição de dois prótons, forma-se a água.