1. Junções estreitas
2. Junções aderentes
3. Desmosmosomas
3. Hemidesmosomas
Células aderem a outras células e à matriz extracelular por meio de proteínas transmembranas de adesão. Destacam-se as integrinas, cadherinas, selectinas e imunoglobulinas. Por vezes, muitas proteínas de adesão unem-se a outras, e a outras proteínas, para formar grandes complexos de adesão macromolecular, geralmente conhecidos como junções celulares. Estes andaimes moleculares são essenciais para manter a integridade de muitos tecidos, tais como tecidos epiteliais, musculares, e nervosos.
As junções celulares são classificadas de acordo com a sua morfologia, o tipo de moléculas de adesão que contêm, as estruturas a que aderem, e as suas interacções com o citoesqueleto. As junções celulares foram inicialmente observadas com microscópios electrónicos e foram nomeadas de acordo com a sua morfologia, mas foram as técnicas de biologia molecular que descobriram a sua complexa organização molecular.
Junções estreitas
Junções estreitas (Figura 1), também conhecidas como zonula ocludens, encontram-se na parte mais apical das células epiteliais, no músculo cardíaco, no edotheium do sistema nervoso central, e nos hepatócitos. Eles fazem contactos tão fortes e apertados entre células adjacentes que o espaço intercelular está quase ocluído.
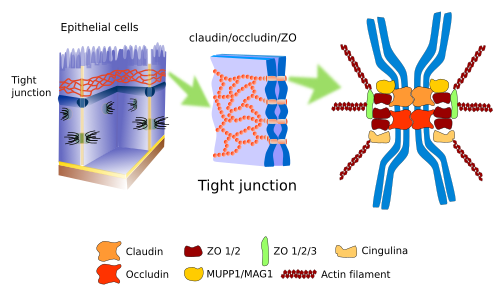
Nas células epiteliais, as junções apertadas funcionam como um fecho de correr em torno do perímetro celular. Para além da forte aderência, as junções apertadas desempenham outras funções. Por exemplo, no epitélio intestinal, as junções apertadas não permitem que as moléculas se difundam através do espaço intercelular entre os enterócitos, e são forçadas a atravessar o enterócito, um caminho muito mais selectivo. Além disso, as junções apertadas mantêm a polaridade das células epiteliais porque formam uma barreira física, de modo a impedir a difusão lateral das moléculas da membrana, tanto de proteínas como de lípidos. É uma barreira física para a difusão lateral na membrana plasmática. Assim, um domínio apical da membrana plasmática é estabelecido com um conjunto particular de moléculas, diferente do domínio laterobasal. Isto é essencial para dirigir adequadamente as moléculas do lúmen intestinal para os vasos sanguíneos: entrar no enterócito através do domínio apical e sair através do domínio basolateral.
As junções estreitas são compostas por mais de 40 tipos de proteínas. As proteínas transmembranas são as claudinas, uma família de proteínas conhecidas como oclusinas, e as proteínas JAM (Junctional Adhesion Molecules). As claudinas são proteínas transmembranas encarregadas de fazer a adesão celular, e entre os pontos de adesão existe um espaço extracelular muito estreito (cerca de 1 nm) que permite aos iões viajarem entre as células. Existem vinte tipos de claudinas que formam passagens com tamanhos diferentes. As células podem alterar a expressão dos tipos de claudinas, regulando assim a permeabilidade através do espaço intercelular. As oclusinas não são realmente necessárias para junções apertadas, mas fornecem estabilidade à junção celular e ajudam a criar uma melhor barreira. As proteínas JAM estabelecem ligações intercelulares, mas parecem ser mais importantes na estabilização do andaime macromolecular. O domínio intracelular destas proteínas está associado a outras moléculas conhecidas como ZO (zonula ocludens), que por sua vez estão ligadas a filamentos de actina e outras proteínas citosólicas. Dependendo do estado de aderência da célula, estas interacções moleculares intracelulares podem desencadear vias de sinalização que afectam a fisiologia celular. Uma observação interessante é que a ocorrência de junções estreitas nas células pode necessitar da presença de junções aderentes.
Junções aderentes
As junções aderentes (zonula adherens) são junções celulares entre células epiteliais. Encontram-se na parte apical da célula, logo abaixo das junções estreitas. Durante o desenvolvimento, as junções aderentes são a primeira junção celular a aparecer em epitélio, antes das junções apertadas. Pensa-se que estão envolvidas em processos morfogenéticos. Como as junções estreitas, as junções aderentes formam uma estrutura semelhante a uma cintura em toda a célula. Os domínios extracelulares de E-Cadherins e nectinas estão envolvidos nos contactos celulares, enquanto que os seus domínios intracelulares estão ligados a várias moléculas como β- y α-catenins, catenin p120 e afadin. Estas proteínas são intermediários entre as moléculas de adesão e os filamentos de actina. Além disso, a β-catenin pode entrar no núcleo e alterar a expressão genética.
As junções dos Aderentes são montadas sequencialmente. Primeiro, são estabelecidas junções mediadas por nectinas, que formam aderências bastante fracas, e depois são recrutadas cadherinas para formar contactos mais fortes e mais estáveis entre células. Em alguns tipos celulares, as junções aderentes iniciam a formação de junções apertadas. As oclusinas, juntamente com o seu andaime de proteínas, são primeiro recrutadas pelas junções aderentes, e as proteínas ZO podem ter um papel relevante neste processo.
Junções apertadas, junções aderentes e desmosomas são necessárias para a integridade da camada epitelial. Mas apenas as junções aderentes são necessárias para os movimentos coordenados de grupos de células epiteliais através da camada epitelial, o que é um movimento relativamente frequente. As aderências de células feitas pelas junções aderentes constroem uma rede ligada de feixes de filamentos de actina a partir de diferentes células que unem uma população celular. Estas células ligadas comportam-se de forma coordenada, por exemplo, para cobrir uma região ferida.
Desmosomas
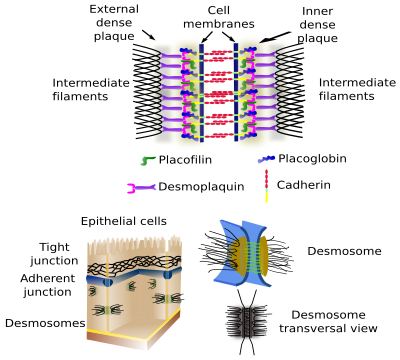
Desmosomas (macula adherens) fazem aderências de células semelhantes às células, como rebites (Figura 2). São muito abundantes em tecidos epiteliais e musculares, mas também podem ser encontrados em praticamente todos os tecidos, por exemplo, no tecido nervoso. Cadherins (desmoglein e desmocollin) são as proteínas de aderência nos desmosomas. Os domínios intracelulares dos cadherins do desmosoma estão associados a filamentos intermediários através de proteínas intermediárias.
4. Hemidesmosomes
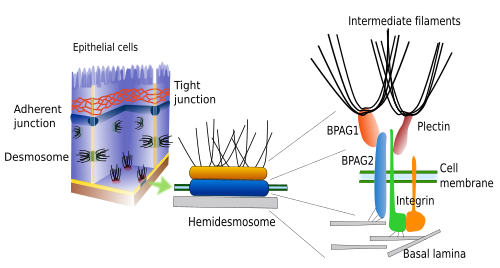
Hemidesmosomes e aderências focais fazem contactos de adesão entre as células e a matriz extracelular (Figura 3). Ambas contêm integrinas como molécula de aderência. Os hemidesmosomas são junções entre as células epiteliais e a lâmina basal (um tipo de matriz extracelular). Intracelularmente, o domínio cistólico destas integrinas está associado a filamentos intermédios. Embora os hemidesmosomas pareçam metade de um desmosoma, são compostos de diferentes moléculas. As aderências focais são menores em tamanho e fazem junções entre células e matriz extracelular por meio de integrinas, que são fixadas intracelularmente a filamentos actínicos.
É comum incluir as junções de fendas dentro do grupo de junções de células. As uniões de fendas são canais moleculares localizados na membrana plasmática que permitem a comunicação directa entre o citoplasma de células adjacentes. Cada canal, conhecido como connexon, é constituído por 6 connexins, e possibilita a difusão de pequenas moléculas entre ambos os citoplasmas. É por isso que as junções de fendas devem ser consideradas como complexos moleculares de comunicação célula-célula em vez de envolvidas na adesão célula-célula.

Bibliografia
Campbell HK, Maiers JL, DeMali KA. (2017). Interacção entre junções apertadas e junções aderentes. Pesquisa de células experimentais 358: 39-44.
Hahn B-S, Labouesse M. (2001). Integridade do tecido: Hemidesmosomas e resistência ao stress. Biologia actual 11:R858-R861.
Huber O. (2003). Estrutura e função das proteínas desmosomais e o seu papel no desenvolvimento e na doença. Ciência da vida celular e molecular. 60:1872-1890.
Ladoux B, Mege RM. (2017). Mecanobiologia dos comportamentos colectivos das células. Revisões da natureza em biologia celular molecular. 18:743-757.
Niessen CM. (2007). Junções estreitas/uniões aderentes: estrutura básica e função. Journal of investigative dermatology. 127:2525-2532.