Oddychanie tlenowe wymaga tlenu (O2) w celu wytworzenia ATP. Chociaż węglowodany, tłuszcze i białka są zużywane jako reaktanty, oddychanie tlenowe jest preferowaną metodą rozkładu pirogronianu w glikolizie i wymaga dostarczenia pirogronianu do mitochondriów w celu jego pełnego utlenienia w cyklu kwasu cytrynowego. Produktami tego procesu są dwutlenek węgla i woda, a przekazana energia jest wykorzystywana do zerwania wiązań w ADP w celu dodania trzeciej grupy fosforanowej, aby utworzyć ATP (adenozynotrójfosforan), poprzez fosforylację na poziomie substratu, NADH i FADH2
| Reakcja uproszczona: | C6H12O6 (s) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + ciepło |
| ΔG = -2880 kJ na mol C6H12O6 |
Ujemne ΔG wskazuje, że reakcja może zachodzić spontanicznie.
Potencjał NADH i FADH2 jest przekształcany w większą ilość ATP poprzez łańcuch transportu elektronów z tlenem i protonami (wodorem) jako „terminalnymi akceptorami elektronów”. Większość ATP wytwarzanego w procesie oddychania tlenowego powstaje w procesie fosforylacji oksydacyjnej. Energia uwolnionego O2 jest wykorzystywana do wytworzenia potencjału chemiosmotycznego poprzez przepompowanie protonów przez błonę. Potencjał ten jest następnie wykorzystywany do napędzania syntazy ATP i wytwarzania ATP z ADP i grupy fosforanowej. Podręczniki biologii często podają, że na jedną utlenioną cząsteczkę glukozy podczas oddychania komórkowego można wytworzyć 38 cząsteczek ATP (2 z glikolizy, 2 z cyklu Krebsa i około 34 z systemu transportu elektronów). Jednak ta maksymalna wydajność nigdy nie jest w pełni osiągnięta z powodu strat spowodowanych nieszczelnymi błonami, jak również kosztem przeniesienia pirogronianu i ADP do macierzy mitochondrialnej, a obecne szacunki wahają się od 29 do 30 ATP na glukozę.
Metabolizm beztlenowy jest do 15 razy bardziej wydajny niż metabolizm beztlenowy (który daje 2 cząsteczki ATP na 1 cząsteczkę glukozy), ponieważ wiązanie podwójne w O2 ma wyższą energię niż inne wiązania podwójne lub pary wiązań pojedynczych w innych wspólnych cząsteczkach w biosferze. Jednak niektóre organizmy beztlenowe, takie jak metanogeny, są w stanie kontynuować oddychanie beztlenowe, uzyskując więcej ATP dzięki wykorzystaniu innych nieorganicznych cząsteczek (nie tlenu) o wysokiej energii jako końcowych akceptorów elektronów w łańcuchu transportu elektronów. Dzielą one początkowy szlak glikolizy, ale metabolizm tlenowy jest kontynuowany przez cykl Krebsa i fosforylację oksydacyjną. Reakcje postglikolityczne zachodzą w mitochondriach w komórkach eukariotycznych i w cytoplazmie w komórkach prokariotycznych.
Glikoliza
Glikoliza to szlak metaboliczny, który zachodzi w cytozolu komórek we wszystkich organizmach żywych. Glikoliza może być dosłownie tłumaczona jako „rozszczepianie cukru” i zachodzi w obecności lub bez obecności tlenu. W warunkach tlenowych proces ten przekształca jedną cząsteczkę glukozy w dwie cząsteczki pirogronianu (kwasu pirogronowego), generując energię w postaci dwóch cząsteczek netto ATP. W rzeczywistości produkowane są cztery cząsteczki ATP na glukozę, jednak dwie są zużywane w fazie przygotowawczej. Wstępna fosforylacja glukozy jest konieczna, aby zwiększyć jej reaktywność (zmniejszyć jej stabilność) w celu rozłożenia jej na dwie cząsteczki pirogronianu przez enzym aldolazę. Podczas fazy zwrotnej glikolizy, cztery grupy fosforanowe są przenoszone na ADP w wyniku fosforylacji na poziomie substratu, aby wytworzyć cztery ATP, a dwie NADH są produkowane, gdy pirogronian jest utleniany. Ogólna reakcja może być wyrażona w ten sposób:
Glukoza + 2 NAD+ + 2 Pi + 2 ADP → 2 pirogronian + 2 H+ + 2 NADH + 2 ATP + 2 H+ + 2 H2O + energia
Zaczynając od glukozy, 1 ATP jest używany do oddania fosforanu do glukozy w celu wytworzenia glukozo-6-fosforanu. Glikogen może być również przekształcony w 6-fosforan glukozy z pomocą fosforylazy glikogenowej. Podczas metabolizmu energetycznego 6-fosforan glukozy przekształca się w 6-fosforan fruktozy. Dodatkowy ATP jest wykorzystywany do fosforylacji fruktozo-6-fosforanu do fruktozo-1,6-bisfosforanu przy udziale fosfofruktokinazy. Fruktozo-1,6-bisfosforan następnie rozdziela się na dwie fosforylowane cząsteczki z trzema łańcuchami węglowymi, które później ulegają degradacji do pirogronianu.
Oksydacyjna dekarboksylacja pirogronianu
Pirogronian jest utleniany do acetylo-CoA i CO2 przez kompleks dehydrogenazy pirogronianowej (PDC). PDC zawiera wiele kopii trzech enzymów i jest zlokalizowany w mitochondriach komórek eukariotycznych oraz w cytozolu prokariotów. W przemianie pirogronianu do acetylo-CoA powstaje jedna cząsteczka NADH i jedna cząsteczka CO2.
Cykl kwasu cytrynowego
Nazywa się go również cyklem Krebsa lub cyklem kwasu trójkarboksylowego. Gdy obecny jest tlen, z cząsteczek pirogronianu powstałych w wyniku glikolizy wytwarzany jest acetylo-CoA. Po utworzeniu acetylo-CoA może nastąpić oddychanie tlenowe lub beztlenowe. Gdy obecny jest tlen, mitochondria przechodzą oddychanie tlenowe, które prowadzi do cyklu Krebsa. Jeśli jednak tlen nie jest obecny, następuje fermentacja cząsteczki pirogronianu. W obecności tlenu, gdy wytwarzany jest acetylo-CoA, cząsteczka ta wchodzi do cyklu kwasu cytrynowego (cykl Krebsa) wewnątrz macierzy mitochondrialnej i jest utleniana do CO2, jednocześnie redukując NAD do NADH. NADH może być wykorzystany przez łańcuch transportu elektronów do wytworzenia kolejnego ATP w ramach fosforylacji oksydacyjnej. Aby w pełni utlenić równowartość jednej cząsteczki glukozy, dwa acetylo-CoA muszą zostać zmetabolizowane przez cykl Krebsa. Podczas tego cyklu powstają dwa niskoenergetyczne produkty odpadowe, H2O i CO2.
Cykl kwasu cytrynowego to 8-stopniowy proces, w którym uczestniczy 18 różnych enzymów i koenzymów. Podczas tego cyklu acetylo-CoA (2 węgle) + oksalooctan (4 węgle) dają cytrynian (6 węgli), który jest przekształcany w bardziej reaktywną formę zwaną izoctanem (6 węgli). Izocyjanian jest modyfikowany do α-ketoglutaranu (5 karbonów), sukcynylo-CoA, bursztynianu, fumaranu, jabłczanu i w końcu do oksalooctanu.
Zyskiem netto z jednego cyklu są 3 NADH i 1 FADH2 jako związki przenoszące wodór (proton plus elektron) oraz 1 wysokoenergetyczny GTP, który może być następnie użyty do produkcji ATP. Tak więc, całkowita wydajność z 1 cząsteczki glukozy (2 cząsteczki pirogronianu) wynosi 6 NADH, 2 FADH2 i 2 ATP.
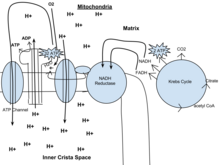
Fosforylacja oksydacyjna
W eukariotach fosforylacja oksydacyjna zachodzi w krysztale mitochondrialnym. Obejmuje ona łańcuch transportu elektronów, który ustanawia gradient protonów (potencjał chemiosmotyczny) przez granicę błony wewnętrznej poprzez utlenianie NADH wytwarzanego z cyklu Krebsa. ATP jest syntetyzowany przez enzym syntazę ATP, gdy gradient chemiosmotyczny jest wykorzystywany do fosforylacji ADP. Transfer elektronów jest napędzany energią chemiczną egzogennego tlenu i z dodatkiem dwóch protonów powstaje woda.
.