
Dziedziczenie cech dla ludzi oparte jest na modelu dziedziczenia Gregora Mendla. Mendel wywnioskował, że dziedziczenie zależy od dyskretnych jednostek dziedziczenia, zwanych czynnikami lub genami.
Dziedziczenie autosomalne dominująceEdit
Cechy autosomalne są związane z pojedynczym genem na autosomie (chromosomie nie będącym chromosomem płci) – są one nazywane „dominującymi”, ponieważ pojedyncza kopia – odziedziczona od któregokolwiek z rodziców – wystarczy, aby spowodować pojawienie się tej cechy. Często oznacza to, że jedno z rodziców musi również posiadać tę samą cechę, chyba że powstała ona w wyniku mało prawdopodobnej nowej mutacji. Przykładami cech i zaburzeń dziedziczonych autosomalnie dominująco są choroba Huntingtona i achondroplazja.
Dziedziczenie autosomalne recesywneEdit

Autosomalne cechy recesywne to jeden wzór dziedziczenia dla cechy, choroby lub zaburzenia, które są przekazywane przez rodziny. Dla recesywnej cechy lub choroby, aby być wyświetlane dwie kopie cechy lub zaburzenia musi być przedstawiony. Cecha lub gen będzie zlokalizowany na chromosomie nie będącym chromosomem płci. Ponieważ do wystąpienia cechy potrzebne są dwie kopie cechy, wiele osób może nieświadomie być nosicielami choroby. Z ewolucyjnego punktu widzenia, recesywna choroba lub cecha może pozostawać w ukryciu przez kilka pokoleń, zanim ujawni się fenotyp. Przykładami zaburzeń dziedziczonych autosomalnie recesywnie są bielactwo, mukowiscydoza.
Dziedziczenie sprzężone z X i Y
Geny sprzężone z X znajdują się na chromosomie płci X. Geny sprzężone z chromosomem X, podobnie jak geny autosomalne, mają zarówno typy dominujące, jak i recesywne. Zaburzenia recesywne związane z chromosomem X są rzadko spotykane u kobiet i zazwyczaj dotyczą tylko mężczyzn. Dzieje się tak dlatego, że mężczyźni dziedziczą swój chromosom X i wszystkie geny związane z chromosomem X będą dziedziczone od strony matki. Ojcowie przekazują swój chromosom Y tylko synom, więc żadne cechy związane z chromosomem X nie będą dziedziczone z ojca na syna. Mężczyźni nie mogą być nosicielami recesywnych cech sprzężonych z chromosomem X, ponieważ mają tylko jeden chromosom X, więc każda cecha sprzężona z chromosomem X odziedziczona po matce pojawi się.
Kobiety wyrażają zaburzenia sprzężone z chromosomem X, gdy są homozygotyczne dla danego zaburzenia i stają się nosicielkami, gdy są heterozygotyczne. Dziedziczenie dominujące X-linked pokaże ten sam fenotyp jako heterozygoty i homozygoty. Podobnie jak w przypadku dziedziczenia sprzężonego z chromosomem X, brak będzie dziedziczenia męsko-męskiego, co pozwala na odróżnienie tego zaburzenia od cech autosomalnych. Jednym z przykładów cechy sprzężonej z chromosomem X jest zespół Coffina-Lowry’ego, który jest spowodowany mutacją w genie białka rybosomalnego. Mutacja ta powoduje nieprawidłowości szkieletowe, czaszkowo-twarzowe, opóźnienie umysłowe i niski wzrost.
Chromosomy X u kobiet przechodzą proces znany jako inaktywacja X. Inaktywacja chromosomu X ma miejsce, gdy jeden z dwóch chromosomów X u kobiet jest prawie całkowicie unieczynniony. Ważne jest, aby proces ten zachodził, w przeciwnym razie kobieta produkowałaby dwa razy więcej normalnych białek chromosomu X. Mechanizm inaktywacji chromosomu X zachodzi na etapie embrionalnym. W przypadku osób z zaburzeniami takimi jak trisomia X, gdzie genotyp posiada trzy chromosomy X, inaktywacja chromosomu X spowoduje inaktywację wszystkich chromosomów X, aż do momentu, gdy aktywny będzie tylko jeden chromosom X. Mężczyźni z zespołem Klinefeltera, którzy mają dodatkowy chromosom X, również zostaną poddani inaktywacji chromosomu X, aby mieć tylko jeden całkowicie aktywny chromosom X.
Dziedziczenie sprzężone z chromosomem Y występuje, gdy gen, cecha lub zaburzenie jest przenoszone przez chromosom Y. Ponieważ chromosomy Y można znaleźć tylko u mężczyzn, cechy związane z chromosomem Y są przekazywane tylko z ojca na syna. Czynnik determinujący jądro, który znajduje się na chromosomie Y, określa męskość osobników. Poza męskością dziedziczoną w chromosomie Y nie ma innych cech sprzężonych z chromosomem Y.
Analiza rodowodówEdit

Rodowód jest diagramem przedstawiającym relacje przodków i przekazywanie cech genetycznych przez kilka pokoleń w rodzinie. Symbole kwadratowe są prawie zawsze używane do reprezentowania samców, podczas gdy kółka są używane dla samic. Rodowody są wykorzystywane do wykrywania wielu różnych chorób genetycznych. Rodowód może być również wykorzystany do określenia szans rodzica na uzyskanie potomstwa z określoną cechą.
Cztery różne cechy mogą być zidentyfikowane poprzez analizę wykresu rodowodowego: autosomalna dominująca, autosomalna recesywna, sprzężona z chromosomem X lub sprzężona z chromosomem Y. Częściowa penetracja może być pokazana i obliczona na podstawie rodowodów. Penetracja to wyrażona procentowo częstotliwość, z jaką osobniki o danym genotypie przejawiają przynajmniej pewien stopień określonego zmutowanego fenotypu związanego z cechą.
Inbreeding, czyli kojarzenie pomiędzy blisko spokrewnionymi organizmami, może być wyraźnie widoczny na wykresach rodowodowych. Wykresy rodowodowe rodzin królewskich często mają wysoki stopień chowu wsobnego, ponieważ było to zwyczajowe i preferowane dla rodzin królewskich, aby poślubić innego członka rodziny królewskiej. Doradcy genetyczni powszechnie używają rodowodów, aby pomóc parom określić, czy rodzice będą w stanie wyprodukować zdrowe dzieci.
KariotypEdit
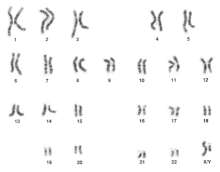
Kariotyp jest bardzo użytecznym narzędziem w cytogenetyce. Kariotyp jest obrazem wszystkich chromosomów w stadium metafazy ułożonych według długości i pozycji centromerowej. Kariotyp może być również przydatny w genetyce klinicznej, ze względu na jego zdolność do diagnozowania zaburzeń genetycznych. Na prawidłowym kariotypie można wykryć aneuploidię poprzez wyraźne zaobserwowanie brakujących lub dodatkowych chromosomów.
Pasmowanie Giemsy, g-banding, kariotypu może być wykorzystane do wykrycia delecji, insercji, duplikacji, inwersji i translokacji. G-banding zabarwi chromosomy jasnymi i ciemnymi pasmami unikalnymi dla każdego chromosomu. FISH, fluorescencyjna hybrydyzacja in situ, może być stosowana do obserwacji delecji, insercji i translokacji. FISH wykorzystuje sondy fluorescencyjne do wiązania się z określonymi sekwencjami chromosomów, które powodują fluorescencję chromosomów w unikalnym kolorze.