Można rozróżnić dwie ogólne klasy zmian ewolucyjnych: mikroewolucję (zmiana poniżej poziomu gatunku) i makroewolucję (zmiana powyżej poziomu gatunku).
Ekolodzy populacji, biolodzy zachowawczy i ekolodzy behawioralni są najbardziej bezpośrednio zainteresowani procesami mikroewolucyjnymi. Obejmują one przesunięcia w wartościach i częstotliwościach poszczególnych cech wśród członków populacji, często z powodu procesów ekologicznych, takich jak przemieszczanie się organizmów i zmieniające się warunki środowiskowe, a także interakcje z członkami różnych gatunków (np. interakcje drapieżnik-pasożyt, żywiciel-pasożyt, konkurencja) lub tego samego gatunku (np. dobór płciowy, konkurencja). Procesy te mogą, ale nie muszą, prowadzić do powstawania nowych gatunków w czasie, lecz zamiast tego powodują fluktuacje częstotliwości cech w populacjach śledzących ciągle zmieniające się presje selekcyjne (Thompson 1998). Ponieważ niektóre procesy mikroewolucyjne mogą zachodzić w ciągu zaledwie kilku pokoleń, często można je obserwować w przyrodzie lub w laboratorium.
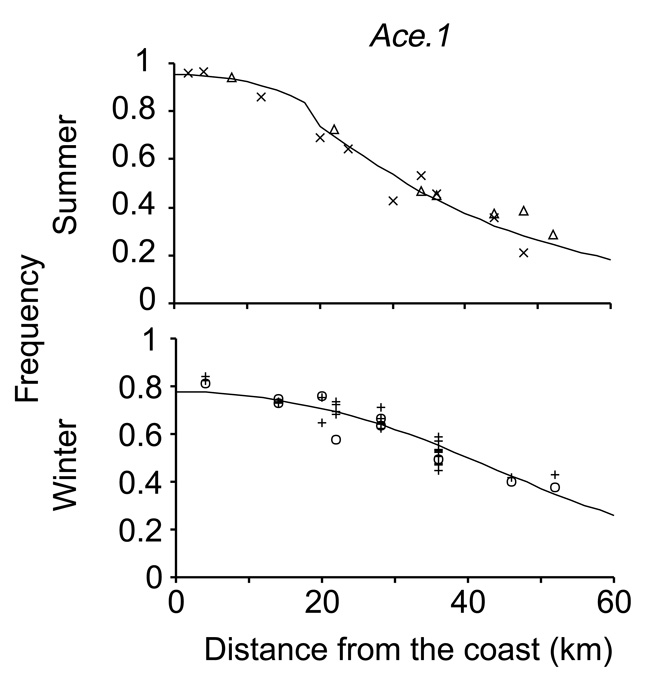
Właściwą ilustracją mikroewolucji w działaniu jest dobrze udokumentowana tendencja owadów do szybkiego rozwijania odporności na pestycydy (Gassmann et al. 2009). Na przykład, podczas lata w południowej Francji, pestycydy są stosowane do zwalczania komarów Culex od wybrzeża Morza Śródziemnego do około 20 km w głąb lądu. Niektóre geny komarów nadają odporność na pestycydy, ale są kosztowne w przypadku braku pestycydów (Rysunek 2); częstotliwość występowania genu odporności na pestycydy wzrasta w okresie letnim na obszarach, gdzie opryski są powszechne, ale nie wzrasta na obszarach, gdzie opryski nie są praktykowane. (Lenormand et al. 1999).
Zazwyczaj zmiany makroewolucyjne nie mogą być typowo obserwowane bezpośrednio ze względu na duże skale czasowe zazwyczaj zaangażowane, chociaż wiele przypadków zmian makroewolucyjnych zostało zaobserwowanych w laboratorium (Rice & Hostert 1993). Zamiast tego, badania makroewolucji polegają na wnioskowaniu na podstawie dowodów kopalnych, rekonstrukcji filogenetycznej i ekstrapolacji z wzorców mikroewolucyjnych. Często badania makroewolucyjne koncentrują się na specjacji: procesie, w którym grupy wcześniej krzyżujących się organizmów stają się niezdolne (lub niechętne) do udanego kojarzenia się ze sobą i produkowania płodnego potomstwa.
Ekolodzy mogą być zainteresowani makroewolucją jako środkiem do wnioskowania na temat współczesnych kwestii ekologicznych. Naukowcy zainteresowani w modelowaniu skutków obecnych zmian klimatycznych, na przykład, może połączyć prehistorycznych danych klimatologicznych z kopalnych pochodzących wzorców specjacji i wymierania, aby zrozumieć, jak współczesne gatunki zwierząt i roślin są faring dziś i jak będą one fare w przyszłości. Na przykład, wiele morskich bezkręgowców (np. koralowce, ślimaki, małże) buduje swoje muszle przy użyciu węglanu wapnia pozyskiwanego z wody oceanicznej. W miarę gromadzenia się antropogenicznego CO2 w atmosferze, znaczna jego część rozpuszcza się w oceanie, uwalniając w tym procesie wolne jony wodorowe i obniżając w ten sposób pH oceanu. Między innymi, to zakwaszenie oceanu zmniejsza ilość węglanu dostępnego dla bezkręgowców morskich tworzących skorupy, które polegają na nim dla swoich wapniowo-węglanowych muszli, co utrudnia im tworzenie i utrzymywanie swoich muszli.
Poprzez połączenie danych oceanicznych pH od setek milionów lat temu z zapisów kopalnych foramifera (shell-making morskich bezkręgowców), Zachos et al. (2005) pokazują skutki, że zakwaszenie oceanu miały na dywersyfikację i wymieranie przeszłości morskiej fauny bezkręgowców. Na podstawie tych danych można modelować aktualne wzorce zakwaszenia oceanu i zacząć przewidywać jego wpływ na obecne i przyszłe zwierzęta morskie (np. Orr et al. 2005).