1. Nauwe juncties
2. Adherente juncties
3. Desmosomen
3. Hemidesmosomen
Cellen hechten zich aan andere cellen en aan de extracellulaire matrix door middel van adhesie transmembraaneiwitten. Integrines, cadherines, selectines en immunoglobulines springen eruit. Soms verbinden vele adhesie-eiwitten zich met elkaar en met andere eiwitten tot grote macromoleculaire adhesiecomplexen, die algemeen bekend staan als celjuncties. Deze moleculaire steigers zijn essentieel om de integriteit van veel weefsels, zoals epitheelweefsel, spierweefsel en zenuwweefsel, in stand te houden.
Celjuncties worden geclassificeerd op basis van hun morfologie, het type adhesiemoleculen dat zij bevatten, de structuren waaraan zij zich hechten, en hun interacties met het cytoskelet. Celjuncties werden voor het eerst waargenomen met elektronenmicroscopen en kregen een naam op basis van hun morfologie, maar het waren de moleculair-biologische technieken die hun complexe moleculaire organisatie blootlegden.
Tight junctions
Tight junctions (figuur 1), ook bekend als zonula occludens, worden aangetroffen in het meer apicale deel van de epitheelcellen, in de hartspier, in het edotheium van het centrale zenuwstelsel, en in hepatocyten. Zij maken zo’n sterke en hechte contacten tussen aangrenzende cellen dat de intercellulaire ruimte bijna wordt afgesloten.
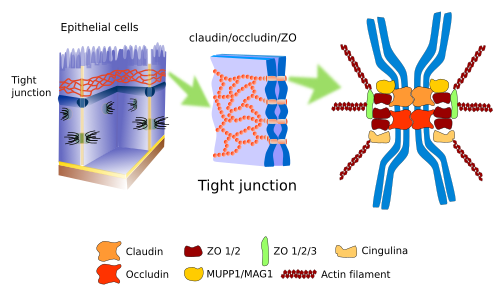
In de epitheelcellen werken tight junctions als een rits rond de omtrek van de cel. Naast de sterke adhesie vervullen tight junctions ook andere functies. In het darmepitheel bijvoorbeeld laten tight junctions niet toe dat moleculen diffunderen door de intercellulaire ruimte tussen enterocyten, en worden ze gedwongen de enterocyt te passeren, een veel selectievere weg. Bovendien houden tight junctions de polariteit van de epitheelcellen in stand omdat zij een fysieke barrière vormen, zodat zij laterale diffusie van membraanmoleculen, zowel eiwitten als lipiden, voorkomen. Zij vormen een fysische barrière voor laterale diffusie in het plasmamembraan. Zo ontstaat een apicaal plasmamembraandomein met een bepaalde set moleculen, verschillend van het laterobasale domein. Dit is essentieel om moleculen vanuit het darmlumen op de juiste wijze naar de bloedvaten te geleiden: de enterocyt binnenkomen via het apicale domein en verlaten via het basolaterale domein.
Tight junctions zijn samengesteld uit meer dan 40 soorten eiwitten. Transmembraan-eiwitten zijn de claudines, een familie van eiwitten die bekend staan als occludines, en de JAM-eiwitten (Junctional Adhesion Molecules). Claudines zijn transmembraaneiwitten die verantwoordelijk zijn voor de cel-cel-adhesie, en tussen de adhesiepunten is er een zeer nauwe extracellulaire ruimte (ongeveer 1 nm) die ionen tussen de cellen laat bewegen. Er zijn twintig soorten claudines die doorgangen van verschillende grootte vormen. Cellen kunnen de expressie van claudinetypes veranderen en zo de permeabiliteit door de intercellulaire ruimte regelen. Occludines zijn niet echt noodzakelijk voor tight junctions, maar zij geven stabiliteit aan de celverbinding en helpen een betere barrière te vormen. JAM-eiwitten brengen intercellulaire verbindingen tot stand, maar zij lijken belangrijker te zijn voor de stabilisatie van de macromoleculaire scaffold. Het intracellulaire domein van deze eiwitten is geassocieerd met andere moleculen, bekend als ZO (zonula occludens), die op hun beurt verbonden zijn met actinefilamenten en andere cytosolische eiwitten. Afhankelijk van de adhesietoestand van de cel kunnen deze intracellulaire moleculaire interacties signaalwegen op gang brengen die de celfysiologie beïnvloeden. Een interessante waarneming is dat voor het ontstaan van tight junction in cellen de aanwezigheid van adherente junctions nodig kan zijn.
Adherente juncties
Adherente juncties (zonula adherens) zijn celjuncties tussen epitheelcellen. Ze bevinden zich in het apicale deel van de cel, net onder de tight junctions. Tijdens de ontwikkeling zijn adherente juncties de eerste cel junctie die verschijnen in epithelia, eerder dan tight junctions. Men denkt dat zij betrokken zijn bij morfogenetische processen. Net als tight junctions vormen adherente juncties een gordelachtige structuur rondom de cel. De extracellulaire domeinen van E-Cadherines en nectines zijn betrokken bij de cel-cel contacten, terwijl hun intracellulaire domeinen gekoppeld zijn aan verschillende moleculen zoals β- y α-catenines, catenine p120 en afadin. Deze eiwitten zijn intermediairs tussen adhesiemoleculen en actinefilamenten. Bovendien kan β-catenine de kern binnendringen en de genexpressie veranderen.
Adherens junctions worden sequentieel geassembleerd. Eerst worden door nectines gemedieerde verbindingen tot stand gebracht, die tamelijk zwakke adhesies vormen, en vervolgens worden cadherines gerekruteerd om sterkere en stabielere contacten tussen cellen te vormen. In sommige celtypes initiëren adherente juncties de vorming van tight junctions. Occludines worden, samen met hun eiwitscaffold, eerst gerekruteerd door de adherente juncties, en ZO eiwitten hebben mogelijk een relevante rol in dit proces.
Strakke juncties, adherente juncties en desmosomen zijn nodig voor de integriteit van de epitheliale laag. Maar alleen de adherente juncties zijn nodig voor de gecoördineerde bewegingen van groepen epitheelcellen door de epitheellaag, wat een relatief frequente beweging is. De cel-cel adhesies gemaakt door adherente juncties bouwen een verbonden net van actine filament bundels van verschillende cellen die een celpopulatie samenvoegen. Deze verbonden cellen gedragen zich gecoördineerd, bijvoorbeeld om een gewond gebied te bedekken.
Desmosomen
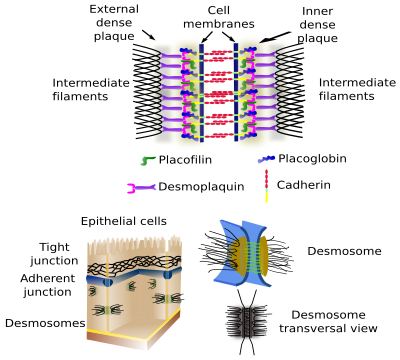
Desmosomen (macula adherens) maken vlekvormige cel-celverklevingen, als klinknagels (figuur 2). Ze zijn zeer talrijk in epitheel- en spierweefsels, maar kunnen ook in vrijwel elk weefsel worden aangetroffen, bijvoorbeeld in het zenuwweefsel. Cadherines (desmogleïne en desmocolline) zijn de adhesie-eiwitten in desmosomen. De intracellulaire domeinen van desmosoom-cadherines zijn via intermediaire eiwitten verbonden met intermediaire filamenten.
4. Hemidesmosomen
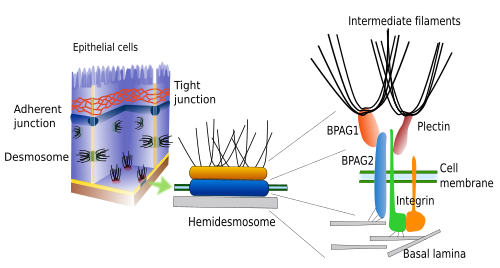
Hemidesmosomen en focale adhesies maken adhesiecontacten tussen cellen en de extracellulaire matrix (figuur 3). Beide bevatten integrines als adhesiemolecuul. Hemidesmosomen zijn knooppunten tussen epiteliale cellen en de basale lamina (een soort extracellulaire matrix). Intracellulair zijn de cystolische domeinen van deze integrinen geassocieerd met intermediaire filamenten. Hoewel hemidesmosomen lijken op de ene helft van een desmosoom, zijn zij samengesteld uit verschillende moleculen. Focal adhesions zijn kleiner van omvang en maken verbindingen tussen cellen en de extracellulaire matrix door middel van integrines, die intracellulair aan actinefilamenten zijn gehecht.
Het is gebruikelijk om gap junctions op te nemen in de groep van celjuncties. Gap junctions zijn moleculaire kanalen in het plasmamembraan die directe communicatie tussen het cytoplasma van aangrenzende cellen mogelijk maken. Elk kanaal, connexon genaamd, bestaat uit 6 connexines en maakt de diffusie van kleine moleculen tussen beide cytoplasma’s mogelijk. Daarom moeten gap junctions worden beschouwd als moleculaire complexen voor cel-cel-communicatie in plaats van als complexen die betrokken zijn bij cel-cel-adhesie.

Bibliografie
Campbell HK, Maiers JL, DeMali KA. (2017). Wisselwerking tussen tight junctions en adherente junctions. Experimenteel celonderzoek 358: 39-44.
Hahn B-S, Labouesse M. (2001). Weefselintegriteit: Hemidesmosomen en weerstand tegen stress. Current biology 11:R858-R861.
Huber O. (2003). Structuur en functie van desmosomale eiwitten en hun rol in ontwikkeling en ziekte. Cell and molecular life science. 60:1872-1890.
Ladoux B, Mege RM. (2017). Mechanobiologie van collectieve celgedragingen. Nature reviews in moleculaire celbiologie. 18:743-757.
Niessen CM. (2007). Tight junctions/adherente juncties: basisstructuur en functie. Tijdschrift voor onderzoekende dermatologie. 127:2525-2532.