進化の変化には、ミクロ進化(種のレベル以下の変化)とマクロ進化(種のレベル以上の変化)の2つの一般的なクラスに分けることができます。
個体群生態学者、保全生物学者、行動生態学者は、ミクロ進化のプロセスに最も直接的に関心を持っています。 これは、生物の移動や環境条件の変化などの生態学的プロセスや、異なる種や同じ種の生物との相互作用(捕食者-被捕食者、宿主-寄生虫の相互作用、競争など)に起因することが多い。 これらのプロセスは、時間の経過とともに新種の形成につながる可能性はあるが、必ずしもそうではなく、刻々と変化する選択圧に追随して集団内の形質の頻度が変動する結果となる(Thompson 1998)。 微小進化プロセスの中には、わずか数世代の間に起こるものもあるため、自然界や実験室で観察することができます。
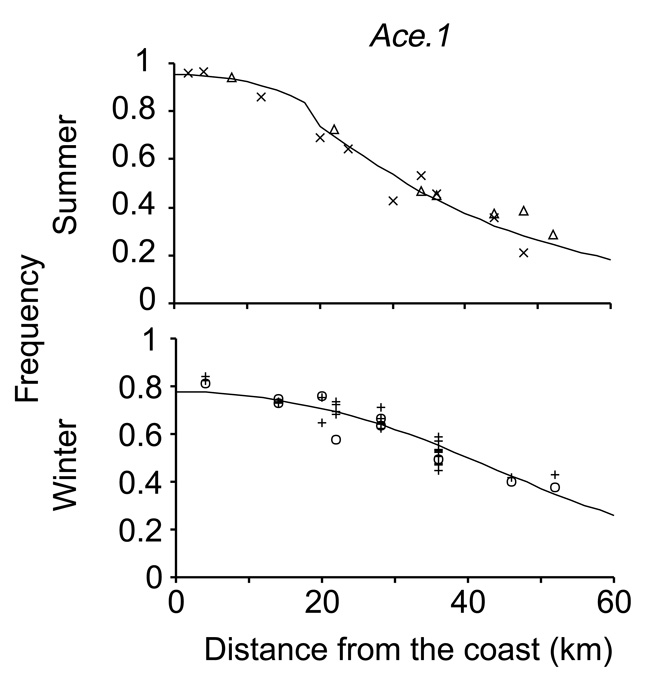
マイクロエボリューションの例としては、昆虫が殺虫剤に対して急速に耐性を獲得する傾向がよく知られています (Gassmann et al. 2009)。 例えば、夏の南フランスでは、地中海沿岸から約20km内陸部まで、蚊を駆除するために農薬が散布されています。 ある種の蚊の遺伝子は、農薬に対する抵抗性をもたらすが、農薬がない場合にはコストがかかる(図2)。農薬抵抗性遺伝子の頻度は、農薬散布が一般的な地域では夏の間に増加するが、散布が行われていない地域では増加しない。 (Lenormand et al. 1999).
マクロ進化の変化は、実験室では多くの例が観察されていますが、一般的には大きな時間スケールを伴うため、直接観察することはできません(Rice & Hostert 1993)。 その代わり、マクロ進化の研究では、化石の証拠からの推論、系統樹の再構築、ミクロ進化のパターンからの外挿に頼る傾向があります。 マクロ進化の研究では、種分化に焦点が当てられることが多くあります。種分化とは、それまで交配していた生物のグループが、お互いにうまく交尾できなくなり、肥沃な子孫を残すことができなくなるプロセスです。 例えば、現在の気候変動の影響をモデル化することに関心のある科学者は、先史時代の気候データと化石由来の種分化や絶滅のパターンを組み合わせることで、現代の動物や植物の種が現在どのような状況にあり、将来どのようになるかを理解することができます。 例えば、多くの海洋無脊椎動物(サンゴ、カタツムリ、アサリなど)は、海水から採取した炭酸カルシウムで殻を作っています。 人為的なCO2が大気中に蓄積されると、そのかなりの部分が海に溶け出し、その過程で遊離水素イオンが放出され、海のpHが低下します。 このような海洋酸性化により、炭酸カルシウムを必要とする海洋無脊椎動物が利用できる炭酸塩の量が減少し、殻の形成や維持が困難になります。
Zachosら(2005)は、数億年前の海洋pHデータと、有孔虫(殻を作る海棲無脊椎動物)の化石記録を組み合わせることで、海洋酸性化が過去の海棲無脊椎動物の多様化と絶滅に与えた影響を示しました。 これらのデータから、現在の海洋酸性化のパターンをモデル化し、現在および将来の海洋動物への影響を予測することができます(例:Olr et al.