La respirazione aerobica richiede ossigeno (O2) per creare ATP. Anche se i carboidrati, i grassi e le proteine sono consumati come reagenti, la respirazione aerobica è il metodo preferito di scissione del piruvato nella glicolisi, e richiede il piruvato ai mitocondri per essere completamente ossidato dal ciclo dell’acido citrico. I prodotti di questo processo sono anidride carbonica e acqua, e l’energia trasferita viene utilizzata per rompere i legami nell’ADP per aggiungere un terzo gruppo fosfato per formare ATP (adenosina trifosfato), mediante fosforilazione a livello di substrato, NADH e FADH2
| Reazione semplificata: | C6H12O6 (s) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + calore |
| ΔG = -2880 kJ per mol di C6H12O6 |
Il ΔG negativo indica che la reazione può avvenire spontaneamente.
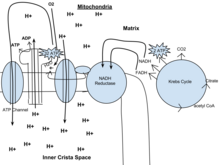
Il potenziale di NADH e FADH2 è convertito in più ATP attraverso una catena di trasporto di elettroni con ossigeno e protoni (idrogeno) come “accettori terminali di elettroni”. La maggior parte dell’ATP prodotta dalla respirazione cellulare aerobica è fatta dalla fosforilazione ossidativa. L’energia dell’O2 rilasciata è usata per creare un potenziale chemiosmotico pompando protoni attraverso una membrana. Questo potenziale è poi usato per guidare l’ATP sintasi e produrre ATP da ADP e un gruppo fosfato. I libri di testo di biologia spesso affermano che 38 molecole di ATP possono essere prodotte per ogni molecola di glucosio ossidato durante la respirazione cellulare (2 dalla glicolisi, 2 dal ciclo di Krebs, e circa 34 dal sistema di trasporto degli elettroni). Tuttavia, questo rendimento massimo non è mai raggiunto del tutto a causa delle perdite dovute alle membrane che perdono e al costo di spostare piruvato e ADP nella matrice mitocondriale, e le stime attuali si aggirano intorno a 29-30 ATP per glucosio.
Il metabolismo aerobico è fino a 15 volte più efficiente del metabolismo anaerobico (che produce 2 molecole di ATP per 1 molecola di glucosio) perché il doppio legame in O2 è di energia superiore ad altri doppi legami o coppie di legami singoli in altre molecole comuni nella biosfera. Tuttavia, alcuni organismi anaerobici, come i metanogeni, sono in grado di continuare con la respirazione anaerobica, producendo più ATP utilizzando altre molecole inorganiche (non l’ossigeno) di alta energia come accettori finali di elettroni nella catena di trasporto degli elettroni. Condividono la via iniziale della glicolisi, ma il metabolismo aerobico continua con il ciclo di Krebs e la fosforilazione ossidativa. Le reazioni post-glicolitiche avvengono nei mitocondri nelle cellule eucariotiche e nel citoplasma nelle cellule procariotiche.
Glicolisi
La glicolisi è una via metabolica che ha luogo nel citosol delle cellule di tutti gli organismi viventi. La glicolisi può essere letteralmente tradotta come “scissione dello zucchero”, e avviene con o senza la presenza di ossigeno. In condizioni aerobiche, il processo converte una molecola di glucosio in due molecole di piruvato (acido piruvico), generando energia sotto forma di due molecole nette di ATP. Quattro molecole di ATP per glucosio sono effettivamente prodotte, tuttavia, due sono consumate come parte della fase preparatoria. La fosforilazione iniziale del glucosio è necessaria per aumentare la reattività (diminuire la sua stabilità) affinché la molecola sia scissa in due molecole di piruvato dall’enzima aldolasi. Durante la fase di pagamento della glicolisi, quattro gruppi fosfato sono trasferiti all’ADP dalla fosforilazione a livello del substrato per produrre quattro ATP, e due NADH sono prodotti quando il piruvato è ossidato. La reazione complessiva può essere espressa in questo modo:
Glucosio + 2 NAD+ + 2 Pi + 2 ADP → 2 piruvato + 2 H+ + 2 NADH + 2 ATP + 2 H+ + 2 H2O + energia
Partendo dal glucosio, 1 ATP viene usato per donare un fosfato al glucosio per produrre glucosio 6-fosfato. Anche il glicogeno può essere convertito in glucosio 6-fosfato con l’aiuto della glicogeno fosforilasi. Durante il metabolismo energetico, il glucosio 6-fosfato diventa fruttosio 6-fosfato. Un ulteriore ATP è usato per fosforilare il fruttosio 6-fosfato in fruttosio 1,6-bisfosfato con l’aiuto della fosfofruttochinasi. Il fruttosio 1,6-bifosfato si divide poi in due molecole fosforilate con tre catene di carbonio che poi si degradano in piruvato.
Decarbossilazione ossidativa del piruvato
Il piruvato è ossidato ad acetil-CoA e CO2 dal complesso piruvato deidrogenasi (PDC). La PDC contiene copie multiple di tre enzimi e si trova nei mitocondri delle cellule eucariotiche e nel citosol dei procarioti. Nella conversione del piruvato in acetil-CoA, si forma una molecola di NADH e una di CO2.
Ciclo dell’acido citrico
Questo è anche chiamato ciclo di Krebs o ciclo dell’acido tricarbossilico. In presenza di ossigeno, l’acetil-CoA viene prodotto dalle molecole di piruvato create dalla glicolisi. Una volta che l’acetil-CoA è formato, la respirazione aerobica o anaerobica può avvenire. Quando l’ossigeno è presente, i mitocondri subiranno la respirazione aerobica che porta al ciclo di Krebs. Tuttavia, se l’ossigeno non è presente, si verifica la fermentazione della molecola di piruvato. In presenza di ossigeno, quando viene prodotto l’acetil-CoA, la molecola entra nel ciclo dell’acido citrico (ciclo di Krebs) all’interno della matrice mitocondriale, e viene ossidata a CO2 mentre allo stesso tempo riduce il NAD a NADH. Il NADH può essere usato dalla catena di trasporto degli elettroni per creare ulteriore ATP come parte della fosforilazione ossidativa. Per ossidare completamente l’equivalente di una molecola di glucosio, due acetil-CoA devono essere metabolizzati dal ciclo di Krebs. Due prodotti di scarto a bassa energia, H2O e CO2, sono creati durante questo ciclo.
Il ciclo dell’acido citrico è un processo in 8 fasi che coinvolge 18 diversi enzimi e coenzimi. Durante il ciclo, acetil-CoA (2 carboni) + ossalacetato (4 carboni) produce citrato (6 carboni), che viene riorganizzato in una forma più reattiva chiamata isocitrato (6 carboni). L’isocitrato viene modificato per diventare α-chetoglutarato (5 carboni), succinil-CoA, succinato, fumarato, malato e, infine, ossalacetato.
Il guadagno netto di un ciclo è di 3 NADH e 1 FADH2 come composti che trasportano idrogeno (protone più elettrone) e 1 GTP ad alta energia, che può essere usato successivamente per produrre ATP. Così, il rendimento totale da 1 molecola di glucosio (2 molecole di piruvato) è 6 NADH, 2 FADH2 e 2 ATP.
Fosforilazione ossidativa
Negli eucarioti, la fosforilazione ossidativa avviene nelle criste mitocondriali. Comprende la catena di trasporto degli elettroni che stabilisce un gradiente protonico (potenziale chemiosmotico) attraverso il confine della membrana interna ossidando il NADH prodotto dal ciclo di Krebs. L’ATP è sintetizzato dall’enzima ATP sintasi quando il gradiente chemiosmotico è usato per guidare la fosforilazione dell’ADP. Il trasferimento di elettroni è guidato dall’energia chimica dell’ossigeno esogeno e, con l’aggiunta di due protoni, si forma l’acqua.