
L’ereditarietà dei tratti umani si basa sul modello di eredità di Gregor Mendel. Mendel dedusse che l’eredità dipende da unità discrete di eredità, chiamate fattori o geni.
Eredità autosomica dominanteModifica
I tratti autosomici sono associati a un singolo gene su un autosoma (cromosoma non sessuale) – sono chiamati “dominanti” perché una singola copia – ereditata da entrambi i genitori – è sufficiente a far apparire questo tratto. Questo spesso significa che anche uno dei genitori deve avere lo stesso tratto, a meno che non sia sorto a causa di un’improbabile nuova mutazione. Esempi di tratti e disturbi autosomici dominanti sono la malattia di Huntington e l’acondroplasia.
Eredità autosomica recessivaModifica

I tratti autosomici recessivi sono un modello di eredità per un tratto, una malattia o un disturbo da trasmettere attraverso le famiglie. Affinché un tratto recessivo o una malattia siano visualizzati, è necessario che siano presenti due copie del tratto o del disturbo. Il tratto o il gene sarà situato su un cromosoma non sessuale. Poiché ci vogliono due copie di un tratto per mostrare un tratto, molte persone possono essere inconsapevolmente portatrici di una malattia. Da una prospettiva evolutiva, una malattia o un tratto recessivo può rimanere nascosto per diverse generazioni prima di mostrare il fenotipo. Esempi di disturbi autosomici recessivi sono l’albinismo e la fibrosi cistica.
Ereditarietà X-linked e Y-linked
I geni X-linked si trovano sul cromosoma X del sesso. I geni X-linked proprio come i geni autosomici hanno sia tipi dominanti che recessivi. I disturbi X-linked recessivi si vedono raramente nelle femmine e di solito colpiscono solo i maschi. Questo perché i maschi ereditano il loro cromosoma X e tutti i geni X-linked saranno ereditati dal lato materno. I padri trasmettono solo il loro cromosoma Y ai loro figli, quindi nessun tratto X-linked sarà ereditato da padre a figlio. Gli uomini non possono essere portatori di tratti X-linked recessivi, in quanto hanno solo un cromosoma X, quindi qualsiasi tratto X-linked ereditato dalla madre si manifesterà.
Le donne esprimono disturbi X-linked quando sono omozigoti per il disturbo e diventano portatrici quando sono eterozigoti. L’eredità dominante X-linked mostrerà lo stesso fenotipo di un eterozigote e di un omozigote. Proprio come l’eredità X-linked, ci sarà una mancanza di eredità da maschio a maschio, che la rende distinguibile dai tratti autosomici. Un esempio di un tratto X-linked è la sindrome di Coffin-Lowry, che è causata da una mutazione nel gene della proteina ribosomiale. Questa mutazione provoca anomalie scheletriche e craniofacciali, ritardo mentale e bassa statura.
I cromosomi X nelle femmine subiscono un processo noto come inattivazione X. L’inattivazione X è quando uno dei due cromosomi X nelle femmine è quasi completamente inattivo. È importante che questo processo avvenga, altrimenti una donna produrrebbe il doppio delle normali proteine del cromosoma X. Il meccanismo di inattivazione dell’X avviene durante la fase embrionale. Per le persone con disturbi come la trisomia X, dove il genotipo ha tre cromosomi X, l’inattivazione dell’X inattiva tutti i cromosomi X fino a quando ci sarà solo un cromosoma X attivo. I maschi con la sindrome di Klinefelter, che hanno un cromosoma X in più, subiranno anch’essi l’inattivazione X per avere un solo cromosoma X completamente attivo.
L’eredità legata all’Y si verifica quando un gene, un tratto o un disturbo viene trasferito attraverso il cromosoma Y. Poiché i cromosomi Y si trovano solo nei maschi, i tratti legati all’Y vengono trasmessi solo da padre a figlio. Il fattore determinante del testicolo, che si trova sul cromosoma Y, determina la mascolinità degli individui. Oltre alla mascolinità ereditata nel cromosoma Y, non ci sono altre caratteristiche legate all’Y.
Analisi dei pedigreeModifica

Un pedigree è un diagramma che mostra le relazioni ancestrali e la trasmissione dei tratti genetici per diverse generazioni in una famiglia. I simboli quadrati sono quasi sempre usati per rappresentare i maschi, mentre i cerchi sono usati per le femmine. I pedigree sono usati per aiutare a rilevare molte malattie genetiche diverse. Un pedigree può anche essere usato per aiutare a determinare le probabilità per un genitore di produrre una prole con un tratto specifico.
Quattro diversi tratti possono essere identificati dall’analisi del grafico del pedigree: autosomico dominante, autosomico recessivo, x-linked, o y-linked. La penetranza parziale può essere mostrata e calcolata dai pedigree. La penetranza è la percentuale di frequenza espressa con cui gli individui di un dato genotipo manifestano almeno un certo grado di uno specifico fenotipo mutante associato a un tratto.
Inbreeding, o accoppiamento tra organismi strettamente imparentati, può essere chiaramente visto sui grafici dei pedigree. I pedigree delle famiglie reali hanno spesso un alto grado di inbreeding, perché era consuetudine e preferibile per i reali sposare un altro membro della famiglia reale. I consulenti genetici usano comunemente i pedigree per aiutare le coppie a determinare se i genitori saranno in grado di produrre figli sani.
CariotipoModifica
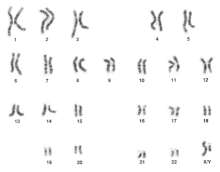
Un cariotipo è uno strumento molto utile in citogenetica. Un cariotipo è un’immagine di tutti i cromosomi in metafase disposti secondo la lunghezza e la posizione dei centromeri. Un cariotipo può essere utile anche nella genetica clinica, grazie alla sua capacità di diagnosticare i disturbi genetici. Su un cariotipo normale, l’aneuploidia può essere rilevata essendo chiaramente in grado di osservare qualsiasi cromosoma mancante o extra.
Il g-banding del cariotipo può essere usato per rilevare delezioni, inserzioni, duplicazioni, inversioni e traslocazioni. Il G-banding colorerà i cromosomi con bande chiare e scure uniche per ogni cromosoma. Una FISH, ibridazione fluorescente in situ, può essere usata per osservare delezioni, inserzioni e traslocazioni. La FISH usa sonde fluorescenti che si legano a specifiche sequenze dei cromosomi e fanno sì che i cromosomi diventino fluorescenti con un colore unico.