La respiration aérobie nécessite de l’oxygène (O2) afin de créer de l’ATP. Bien que les glucides, les graisses et les protéines soient consommés comme réactifs, la respiration aérobie est la méthode privilégiée de décomposition du pyruvate dans la glycolyse, et nécessite que le pyruvate soit acheminé vers les mitochondries afin d’être entièrement oxydé par le cycle de l’acide citrique. Les produits de ce processus sont le dioxyde de carbone et l’eau, et l’énergie transférée est utilisée pour briser les liaisons dans l’ADP afin d’ajouter un troisième groupe phosphate pour former l’ATP (adénosine triphosphate), par phosphorylation au niveau du substrat, le NADH et le FADH2
| Réaction simplifiée : | C6H12O6 (s) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + chaleur |
| ΔG = -2880 kJ par mole de C6H12O6 |
Le ΔG négatif indique que la réaction peut se produire spontanément.
Le potentiel de NADH et de FADH2 est converti en plus d’ATP par une chaîne de transport d’électrons avec l’oxygène et les protons (hydrogène) comme » accepteurs d’électrons terminaux « . La majeure partie de l’ATP produit par la respiration cellulaire aérobie est réalisée par phosphorylation oxydative. L’énergie de l’O2 libérée est utilisée pour créer un potentiel chimiosmotique en pompant des protons à travers une membrane. Ce potentiel est ensuite utilisé pour entraîner l’ATP synthase et produire de l’ATP à partir d’ADP et d’un groupe phosphate. Les manuels de biologie indiquent souvent que 38 molécules d’ATP peuvent être produites par molécule de glucose oxydée pendant la respiration cellulaire (2 par la glycolyse, 2 par le cycle de Krebs et environ 34 par le système de transport des électrons). Cependant, ce rendement maximal n’est jamais tout à fait atteint en raison des pertes dues aux membranes non étanches ainsi que du coût du déplacement du pyruvate et de l’ADP dans la matrice mitochondriale, et les estimations actuelles tournent autour de 29 à 30 ATP par glucose.
Le métabolisme aérobie est jusqu’à 15 fois plus efficace que le métabolisme anaérobie (qui donne 2 molécules d’ATP pour 1 molécule de glucose) parce que la double liaison de l’O2 est d’une énergie plus élevée que d’autres doubles liaisons ou paires de liaisons simples dans d’autres molécules communes de la biosphère. Cependant, certains organismes anaérobies, tels que les méthanogènes, sont capables de poursuivre la respiration anaérobie et de produire plus d’ATP en utilisant d’autres molécules inorganiques (pas l’oxygène) à haute énergie comme accepteurs finaux d’électrons dans la chaîne de transport des électrons. Ils partagent la voie initiale de la glycolyse mais le métabolisme aérobie se poursuit avec le cycle de Krebs et la phosphorylation oxydative. Les réactions post-glycolytiques ont lieu dans les mitochondries des cellules eucaryotes, et dans le cytoplasme des cellules procaryotes.
Glycolyse
La glycolyse est une voie métabolique qui a lieu dans le cytosol des cellules de tous les organismes vivants. La glycolyse peut être littéralement traduite par » division du sucre « , et se produit avec ou sans la présence d’oxygène. Dans des conditions aérobies, le processus convertit une molécule de glucose en deux molécules de pyruvate (acide pyruvique), générant de l’énergie sous la forme de deux molécules nettes d’ATP. Quatre molécules d’ATP par glucose sont effectivement produites, mais deux sont consommées dans le cadre de la phase préparatoire. La phosphorylation initiale du glucose est nécessaire pour augmenter sa réactivité (diminuer sa stabilité) afin que la molécule soit clivée en deux molécules de pyruvate par l’enzyme aldolase. Pendant la phase d’amortissement de la glycolyse, quatre groupes phosphates sont transférés à l’ADP par la phosphorylation au niveau du substrat pour produire quatre ATP, et deux NADH sont produits lorsque le pyruvate est oxydé. La réaction globale peut être exprimée ainsi :
Glucose + 2 NAD+ + 2 Pi + 2 ADP → 2 pyruvate + 2 H+ + 2 NADH + 2 ATP + 2 H+ + 2 H2O + énergie
En partant du glucose, 1 ATP est utilisé pour donner un phosphate au glucose afin de produire du glucose 6-phosphate. Le glycogène peut également être converti en glucose 6-phosphate à l’aide de la glycogène phosphorylase. Au cours du métabolisme énergétique, le glucose 6-phosphate devient du fructose 6-phosphate. Un ATP supplémentaire est utilisé pour phosphoryler le fructose 6-phosphate en fructose 1,6-bisphosphate à l’aide de la phosphofructokinase. Le fructose 1,6-biphosphate se divise ensuite en deux molécules phosphorylées à trois chaînes de carbone qui se dégradent ensuite en pyruvate.
Décarboxylation oxydative du pyruvate
Le pyruvate est oxydé en acétyl-CoA et CO2 par le complexe pyruvate déshydrogénase (PDC). Le PDC contient des copies multiples de trois enzymes et est situé dans les mitochondries des cellules eucaryotes et dans le cytosol des procaryotes. Lors de la conversion du pyruvate en acétyl-CoA, une molécule de NADH et une molécule de CO2 sont formées.
Cycle de l’acide citrique
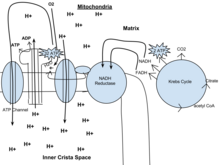
Ce cycle est également appelé cycle de Krebs ou cycle de l’acide tricarboxylique. En présence d’oxygène, l’acétyl-CoA est produit à partir des molécules de pyruvate créées par la glycolyse. Une fois l’acétyl-CoA formé, la respiration aérobie ou anaérobie peut se produire. En présence d’oxygène, la mitochondrie subit une respiration aérobie qui aboutit au cycle de Krebs. En revanche, en l’absence d’oxygène, la fermentation de la molécule de pyruvate se produit. En présence d’oxygène, lorsque l’acétyl-CoA est produit, la molécule entre alors dans le cycle de l’acide citrique (cycle de Krebs) à l’intérieur de la matrice mitochondriale, et est oxydée en CO2 tout en réduisant le NAD en NADH. Le NADH peut être utilisé par la chaîne de transport d’électrons pour créer de l’ATP supplémentaire dans le cadre de la phosphorylation oxydative. Pour oxyder complètement l’équivalent d’une molécule de glucose, deux acétyl-CoA doivent être métabolisés par le cycle de Krebs. Deux déchets à faible énergie, H2O et CO2, sont créés au cours de ce cycle.
Le cycle de l’acide citrique est un processus en 8 étapes impliquant 18 enzymes et co-enzymes différents. Au cours du cycle, acétyl-CoA (2 carbones) + oxaloacétate (4 carbones) donne du citrate (6 carbones), qui est réarrangé en une forme plus réactive appelée isocitrate (6 carbones). L’isocitrate est modifié pour devenir de l’α-cétoglutarate (5 carbones), du succinyl-CoA, du succinate, du fumarate, du malate et, finalement, de l’oxaloacétate.
Le gain net d’un cycle est de 3 NADH et 1 FADH2 en tant que composés porteurs d’hydrogène (proton plus électron) et 1 GTP à haute énergie, qui peut ensuite être utilisé pour produire de l’ATP. Ainsi, le rendement total d’une molécule de glucose (2 molécules de pyruvate) est de 6 NADH, 2 FADH2 et 2 ATP.
Phosphorylation oxydative
Chez les eucaryotes, la phosphorylation oxydative a lieu dans les cristaux mitochondriaux. Elle comprend la chaîne de transport des électrons qui établit un gradient de protons (potentiel chimiosmotique) à travers la limite de la membrane interne en oxydant le NADH produit par le cycle de Krebs. L’ATP est synthétisée par l’enzyme ATP synthase lorsque le gradient chimiosmotique est utilisé pour entraîner la phosphorylation de l’ADP. Le transfert d’électrons est entraîné par l’énergie chimique de l’oxygène exogène et, avec l’ajout de deux protons, l’eau est formée.