1. Les jonctions serrées
2. Les jonctions adhérentes
3. Les desmosomes
3. Les hémidesmosomes
Les cellules adhèrent aux autres cellules et à la matrice extracellulaire grâce à des protéines transmembranaires d’adhésion. Les intégrines, les cadhérines, les sélectines et les immunoglobulines se distinguent. Parfois, de nombreuses protéines d’adhésion se joignent les unes aux autres, et à d’autres protéines, pour former de grands complexes d’adhésion macromoléculaires, généralement appelés jonctions cellulaires. Ces échafaudages moléculaires sont essentiels pour maintenir l’intégrité de nombreux tissus, tels que les tissus épithéliaux, musculaires et nerveux.
Les jonctions cellulaires sont classées en fonction de leur morphologie, du type de molécules d’adhésion qu’elles contiennent, des structures auxquelles elles adhèrent et de leurs interactions avec le cytosquelette. Les jonctions cellulaires ont d’abord été observées avec des microscopes électroniques et ont été nommées en fonction de leur morphologie, mais ce sont les techniques de biologie moléculaire qui ont mis au jour leur organisation moléculaire complexe.
Jonctions serrées
Les jonctions serrées (figure 1), également appelées zonula occludens, se trouvent dans la partie plus apicale des cellules épithéliales, dans le muscle cardiaque, dans l’œdème du système nerveux central et dans les hépatocytes. Ils établissent des contacts si forts et si serrés entre les cellules adjacentes que l’espace intercellulaire est presque occlus.
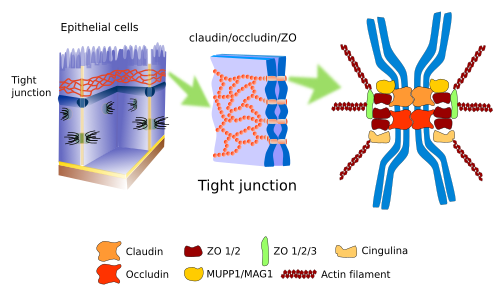
Dans les cellules épithéliales, les jonctions serrées fonctionnent comme une fermeture éclair autour du périmètre cellulaire. Outre la forte adhérence, les jonctions serrées remplissent d’autres fonctions. Par exemple, dans l’épithélium de l’intestin, les jonctions serrées ne permettent pas aux molécules de diffuser dans l’espace intercellulaire entre les entérocytes, et sont obligées de traverser l’entérocyte, une voie beaucoup plus sélective. En outre, les jonctions serrées maintiennent la polarité des cellules épithéliales car elles forment une barrière physique, de sorte qu’elles empêchent la diffusion latérale des molécules membranaires, qu’il s’agisse de protéines ou de lipides. Elles constituent une barrière physique pour la diffusion latérale dans la membrane plasmique. Ainsi, un domaine apical de la membrane plasmique est établi avec un ensemble particulier de molécules, différent du domaine latérobasal. Ceci est essentiel pour diriger correctement les molécules de la lumière de l’intestin vers les vaisseaux sanguins : entrer dans l’entérocyte par le domaine apical et sortir par le domaine latéral.
Les jonctions serrées sont composées de plus de 40 types de protéines. Les protéines transmembranaires sont les claudines, une famille de protéines connues sous le nom d’occludines, et les protéines JAM (Junctional Adhesion Molecules). Les claudines sont des protéines transmembranaires chargées de réaliser l’adhésion cellule-cellule, et entre les points d’adhésion il y a un espace extracellulaire très étroit (environ 1 nm) qui permet aux ions de voyager entre les cellules. Il existe vingt types de claudines qui forment des passages de tailles différentes. Les cellules peuvent modifier l’expression des types de claudines, régulant ainsi la perméabilité de l’espace intercellulaire. Les occludines ne sont pas vraiment nécessaires aux jonctions serrées, mais elles assurent la stabilité de la jonction cellulaire et contribuent à créer une meilleure barrière. Les protéines JAM établissent des connexions intercellulaires, mais elles semblent plus importantes pour stabiliser l’échafaudage macromoléculaire. Le domaine intracellulaire de ces protéines est associé à d’autres molécules appelées ZO (zonula occludens), qui sont à leur tour connectées à des filaments d’actine et à d’autres protéines cytosoliques. En fonction de l’état d’adhésion de la cellule, ces interactions moléculaires intracellulaires peuvent déclencher des voies de signalisation qui affectent la physiologie de la cellule. Une observation intéressante est que la présence de jonctions serrées dans les cellules peut nécessiter la présence de jonctions adhérentes.
Jonctions adhérentes
Les jonctions adhérentes (zonula adherens) sont des jonctions cellulaires entre les cellules épithéliales. Elles se trouvent dans la partie apicale de la cellule, juste en dessous des jonctions serrées. Au cours du développement, les jonctions adhérentes sont la première jonction cellulaire à apparaître dans les épithéliums, avant les jonctions serrées. On pense qu’elles sont impliquées dans les processus morphogénétiques. Comme les jonctions serrées, les jonctions adhérentes forment une structure en forme de ceinture tout autour de la cellule. Les domaines extracellulaires des E-Cadhérines et des nectines sont impliqués dans les contacts cellule-cellule, tandis que leurs domaines intracellulaires sont liés à plusieurs molécules comme les β- y α-caténines, la caténine p120 et l’afadine. Ces protéines sont des intermédiaires entre les molécules d’adhésion et les filaments d’actine. De plus, la β-caténine peut entrer dans le noyau et modifier l’expression des gènes.
Les jonctions d’adhérence sont assemblées de manière séquentielle. D’abord, des jonctions médiées par les nectines sont établies, qui forment des adhésions plutôt faibles, puis les cadhérines sont recrutées pour former des contacts plus forts et plus stables entre les cellules. Dans certains types cellulaires, les jonctions adhérentes initient la formation de jonctions serrées. Les occludines, ainsi que leur échafaudage protéique, sont d’abord recrutées par les jonctions adhérentes, et les protéines ZO pourraient avoir un rôle pertinent dans ce processus.
Les jonctions serrées, les jonctions adhérentes et les desmosomes sont nécessaires à l’intégrité de la couche épithéliale. Mais seules les jonctions adhérentes sont nécessaires pour les mouvements coordonnés de groupes de cellules épithéliales à travers la couche épithéliale, ce qui est un mouvement relativement fréquent. Les adhésions cellule-cellule réalisées par les jonctions adhérentes construisent un réseau connecté de faisceaux de filaments d’actine provenant de cellules différentes qui réunissent une population de cellules. Ces cellules connectées se comportent de manière coordonnée, par exemple pour couvrir une région blessée.
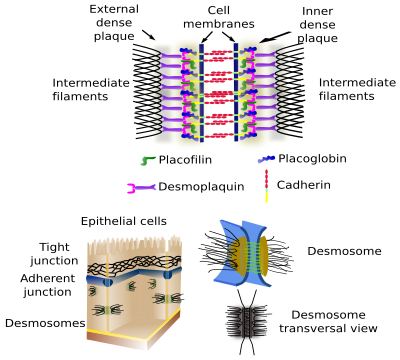
Desmosomes
Les desmosomes (macula adherens) réalisent des adhésions cellule-cellule ponctuelles, comme des rivets (figure 2). Ils sont très abondants dans les tissus épithéliaux et musculaires, mais on peut aussi les trouver dans pratiquement tous les tissus, par exemple dans le tissu nerveux. Les cadhérines (desmoglein et desmocollin) sont les protéines d’adhésion des desmosomes. Les domaines intracellulaires des cadhérines des desmosomes sont associés à des filaments intermédiaires par des protéines intermédiaires.
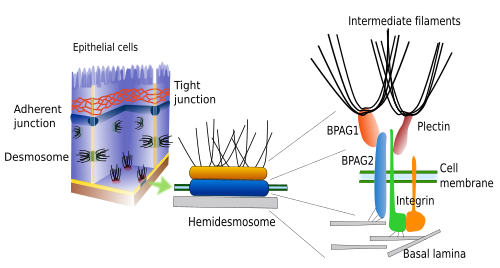
4. Hémidesmosomes
Les hémidesmosomes et les adhésions focales établissent des contacts d’adhésion entre les cellules et la matrice extracellulaire (figure 3). Tous deux contiennent des intégrines comme molécule d’adhésion. Les hémidesmosomes sont des jonctions entre les cellules épithéliales et la lame basale (un type de matrice extracellulaire). Au niveau intracellulaire, le domaine cystolique de ces intégrines est associé à des filaments intermédiaires. Bien que les hémidesmosomes ressemblent à la moitié d’un desmosome, ils sont composés de molécules différentes. Les adhésions focales sont de plus petite taille et font des jonctions entre les cellules et la matrice extracellulaire au moyen d’intégrines, qui sont attachées de manière intracellulaire à des filaments d’actine.
Il est courant d’inclure les jonctions gap dans le groupe des jonctions cellulaires. Les jonctions gap sont des canaux moléculaires situés dans la membrane plasmique qui permettent la communication directe entre le cytoplasme de cellules adjacentes. Chaque canal, appelé connexon, est constitué de 6 connexines, et rend possible la diffusion de petites molécules entre les deux cytoplasmes. C’est pourquoi les jonctions gap doivent être considérées comme des complexes moléculaires de communication entre cellules au lieu d’être impliquées dans l’adhésion cellule-cellule.

Bibliographie
Campbell HK, Maiers JL, DeMali KA. (2017). Interaction entre les jonctions serrées et les jonctions adhérentes. Recherche cellulaire expérimentale 358 : 39-44.
Hahn B-S, Labouesse M. (2001). L’intégrité des tissus : hémidesmosomes et résistance au stress. Current biology 11:R858-R861.
Huber O. (2003). Structure et fonction des protéines desmosomales et leur rôle dans le développement et la maladie. Science de la vie cellulaire et moléculaire. 60:1872-1890.
Ladoux B, Mege RM. (2017). Mécanobiologie des comportements cellulaires collectifs. Nature reviews in molecular cell biology. 18:743-757.
Niessen CM. (2007). Jonctions serrées/jonctions adhérentes : structure et fonction de base. Journal of investigative dermatology. 127:2525-2532.