La respiración aeróbica requiere oxígeno (O2) para crear ATP. Aunque los carbohidratos, las grasas y las proteínas se consumen como reactivos, la respiración aeróbica es el método preferido para la descomposición del piruvato en la glucólisis, y requiere que el piruvato llegue a las mitocondrias para ser completamente oxidado por el ciclo del ácido cítrico. Los productos de este proceso son el dióxido de carbono y el agua, y la energía transferida se utiliza para romper los enlaces en el ADP para añadir un tercer grupo fosfato para formar ATP (trifosfato de adenosina), mediante la fosforilación a nivel de sustrato, NADH y FADH2
| Reacción simplificada: | C6H12O6 (s) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + calor |
| ΔG = -2880 kJ por mol de C6H12O6 |
El ΔG negativo indica que la reacción puede ocurrir espontáneamente.
El potencial de NADH y FADH2 se convierte en más ATP a través de una cadena de transporte de electrones con oxígeno y protones (hidrógeno) como «aceptores terminales de electrones». La mayor parte del ATP producido por la respiración celular aeróbica se realiza mediante la fosforilación oxidativa. La energía del O2 liberado se utiliza para crear un potencial quimiosmótico mediante el bombeo de protones a través de una membrana. Este potencial se utiliza entonces para impulsar la ATP sintasa y producir ATP a partir de ADP y un grupo fosfato. Los libros de texto de biología suelen decir que se pueden producir 38 moléculas de ATP por cada molécula de glucosa oxidada durante la respiración celular (2 de la glucólisis, 2 del ciclo de Krebs y unas 34 del sistema de transporte de electrones). Sin embargo, este rendimiento máximo nunca se alcanza del todo debido a las pérdidas debidas a las membranas con fugas, así como al coste de trasladar el piruvato y el ADP a la matriz mitocondrial, y las estimaciones actuales se sitúan en torno a los 29 o 30 ATP por glucosa.
El metabolismo anaeróbico es hasta 15 veces más eficiente que el anaeróbico (que produce 2 moléculas de ATP por 1 molécula de glucosa) porque el doble enlace del O2 es de mayor energía que otros dobles enlaces o pares de enlaces simples en otras moléculas comunes en la biosfera. Sin embargo, algunos organismos anaeróbicos, como los metanógenos, son capaces de continuar con la respiración anaeróbica, produciendo más ATP al utilizar otras moléculas inorgánicas (no el oxígeno) de alta energía como aceptores finales de electrones en la cadena de transporte de electrones. Comparten la vía inicial de la glucólisis, pero el metabolismo aeróbico continúa con el ciclo de Krebs y la fosforilación oxidativa. Las reacciones postglicolíticas tienen lugar en las mitocondrias en las células eucariotas, y en el citoplasma en las células procariotas.
Glicólisis
La glucólisis es una vía metabólica que tiene lugar en el citosol de las células de todos los organismos vivos. La glucólisis puede traducirse literalmente como «división del azúcar», y se produce con o sin la presencia de oxígeno. En condiciones aeróbicas, el proceso convierte una molécula de glucosa en dos moléculas de piruvato (ácido pirúvico), generando energía en forma de dos moléculas netas de ATP. En realidad se producen cuatro moléculas de ATP por cada glucosa, sin embargo, dos se consumen como parte de la fase preparatoria. La fosforilación inicial de la glucosa es necesaria para aumentar la reactividad (disminuir su estabilidad) para que la molécula sea escindida en dos moléculas de piruvato por la enzima aldolasa. Durante la fase de pago de la glucólisis, se transfieren cuatro grupos fosfato al ADP mediante la fosforilación a nivel de sustrato para producir cuatro ATP, y se producen dos NADH cuando se oxida el piruvato. La reacción global puede expresarse así:
Glucosa + 2 NAD+ + 2 Pi + 2 ADP → 2 piruvato + 2 H+ + 2 NADH + 2 ATP + 2 H+ + 2 H2O + energía
Comenzando con la glucosa, se utiliza 1 ATP para donar un fosfato a la glucosa y producir glucosa 6-fosfato. El glucógeno también puede convertirse en glucosa 6-fosfato con la ayuda de la glucógeno fosforilasa. Durante el metabolismo energético, la glucosa 6-fosfato se convierte en fructosa 6-fosfato. Se utiliza un ATP adicional para fosforilar la fructosa 6-fosfato en fructosa 1,6-bifosfato con la ayuda de la fosfofructoquinasa. A continuación, la fructosa 1,6-bifosfato se divide en dos moléculas fosforiladas con tres cadenas de carbono que posteriormente se degradan en piruvato.
Descarboxilación oxidativa del piruvato
El piruvato es oxidado a acetil-CoA y CO2 por el complejo piruvato deshidrogenasa (PDC). El PDC contiene múltiples copias de tres enzimas y se localiza en las mitocondrias de las células eucariotas y en el citosol de las procariotas. En la conversión de piruvato en acetil-CoA se forma una molécula de NADH y una de CO2.
Ciclo del ácido cítrico
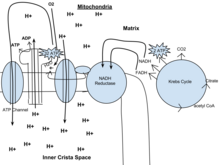
También se denomina ciclo de Krebs o ciclo del ácido tricarboxílico. Cuando hay oxígeno, se produce acetil-CoA a partir de las moléculas de piruvato creadas a partir de la glucólisis. Una vez formado el acetil-CoA, puede producirse la respiración aeróbica o anaeróbica. Cuando el oxígeno está presente, las mitocondrias realizan la respiración aeróbica que conduce al ciclo de Krebs. Sin embargo, si el oxígeno no está presente, se producirá la fermentación de la molécula de piruvato. En presencia de oxígeno, cuando se produce acetil-CoA, la molécula entra entonces en el ciclo del ácido cítrico (ciclo de Krebs) dentro de la matriz mitocondrial, y se oxida a CO2 mientras que al mismo tiempo se reduce el NAD a NADH. El NADH puede ser utilizado por la cadena de transporte de electrones para crear más ATP como parte de la fosforilación oxidativa. Para oxidar completamente el equivalente a una molécula de glucosa, deben metabolizarse dos acetil-CoA mediante el ciclo de Krebs. Durante este ciclo se crean dos productos de desecho de baja energía, H2O y CO2.
El ciclo del ácido cítrico es un proceso de 8 pasos en el que intervienen 18 enzimas y coenzimas diferentes. Durante el ciclo, el acetil-CoA (2 carbonos) + el oxaloacetato (4 carbonos) produce citrato (6 carbonos), que se reordena a una forma más reactiva llamada isocitrato (6 carbonos). El isocitrato se modifica para convertirse en α-cetoglutarato (5 carbonos), succinil-CoA, succinato, fumarato, malato y, finalmente, oxaloacetato.
La ganancia neta de un ciclo es de 3 NADH y 1 FADH2 como compuestos portadores de hidrógeno (protones más electrones) y 1 GTP de alta energía, que puede utilizarse posteriormente para producir ATP. Así, el rendimiento total de 1 molécula de glucosa (2 moléculas de piruvato) es de 6 NADH, 2 FADH2 y 2 ATP.
Fosforilación oxidativa
En los eucariotas, la fosforilación oxidativa se produce en las cristas mitocondriales. Comprende la cadena de transporte de electrones que establece un gradiente de protones (potencial quimiosmótico) a través del límite de la membrana interna mediante la oxidación del NADH producido en el ciclo de Krebs. El ATP es sintetizado por la enzima ATP sintasa cuando el gradiente quimiosmótico se utiliza para impulsar la fosforilación del ADP. La transferencia de electrones es impulsada por la energía química del oxígeno exógeno y, con la adición de dos protones, se forma agua.